
Critique of “Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period” — Part 3: Estimating reproduction numbers
Want to share your content on R-bloggers? click here if you have a blog, or here if you don't.
This is the third in a series of posts (previous posts: Part 1, Part 2) in which I look at the following paper:
Kissler, Tedijanto, Goldstein, Grad, and Lipsitch, Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period, Science, vol. 368, pp. 860-868, 22 May 2020 (released online 14 April 2020). The paper is also available here, with supplemental materials here.
In this post, I’ll look at how the authors estimate the reproduction numbers (R) over time for the four common cold coronavirus, using the proxies for incidence that I discussed in Part 2. These estimates for R are used to model immunity and cross-immunity for these viruses, and the seasonal effects on their transmission. These modelling results inform the later parts of the paper, in which they consider various scenarios for future transmission of SARS-CoV-2 (the coronavirus responsible for COVID-19), whose characteristics may perhaps resemble those of these other coronaviruses.
I will be using the code (written in R) available here, with GPLv2 licence, which I wrote to replicate the results in the paper, and which allows me to more easily produce plots to help understand issues with the methods, and to try out alternative methods that may work better, than the code provided by the authors (which I discussed in Part 1).
Reproduction numbers and generative interval distributions
The reproduction number, R, for a virus is the average number of people that a person infected with the virus will go on to infect. A more detailed description of transmission is provided by the generation interval distribution, the probability distribution for the time interval, a, from a person being infected to them infecting another person, denoted by g(a). The expected number of people infected at time t2 by a particular person infected at time t1 is Rg(t2-t1).
Here, I have assumed that time is an integer, measuring days, so g(a) is a discrete distribution over positive integers. It’s common in the epidemiology literature to treat time as continuous, but this would seem to make little sense unless one is modelling the likely quite complex details of daily cycles in viral physiology and human behaviour, which would lead to complex variation in R and g within each day. As far as I can tell, nobody does this.
It’s expected that R will change over time, both because the fraction of the population that is immune to infection may change, and because environmental influences on transmission of the virus may change, perhaps as the season changes, or as people alter their behaviour to avoid infection. It may also be that g changes with time, but handling that seems to be sufficiently difficult that the possibility is typically ignored.
When R is changing, there are at least two natural ways of defining its value at a given time. One is the average number of people that a person infected at time u will go on to infect, which I (and Kissler, et al.) denote by Ru. Looking backwards gives another definition, which I will denote by Rt — the number of people who become infected at time t divided by the number of people infected before time t, weighted by the probability of them transmitting the virus to time t, according to the generation interval distribution, g.
Kissler, et al. reference a paper by Wallinga and Lipsitch which gives formulas for Rt and Ru (their equations 4.1 and 4.2). For discrete generative interval distributions, these can be written as Rt = b(t) / ∑a b(t-a)g(a) and Ru = ∑t g(t-u)Rt, where b(t) is the number of people who became infected at time t. The sum over a here is from 1 to infinity; the sum over t is from u+1 to infinity.
The Kissler, et al. paper estimates Ru and then tries to model these estimates. Although Ru may seem like a simpler, more natural quantity than Rt, I think that Rt is the better target for modelling. Ru looks into the future — if some intervention, such as a ban on large gatherings, is introduced at time T, this will change Rt only for t≥T. but it will change Ru even at times u
I will look at estimates for both Rt and Ru. The estimates for Rt are needed in any case in order to produce the estimates for Ru. I will also look at the 21-day averaged estimates for Ru that were used by Kissler, et al.
Interpolating to produce daily incidence proxies
The formulas for Rt and Ru above require daily data on new infections, b(t). But the incidence proxies that are used by Kissler, et al., which I discussed in Part 2 of this series, are aggregated over weeks.
As discussed below, a suitable generative interval distribution for these coronaviruses puts significant probability on intervals of only a few days, so switching to a discrete-time representation in terms of weeks would substantially distort the results. Instead, Kissler, et al. interpolate the weekly incidence proxies to daily values. This is done by a procedure described in the supplementary material of a paper by te Beest, et al., as follows:
- The weekly incidence proxy for a virus is used to produce a weekly proxy for cumulative incidence.
- An interpolating spline is fit to the weekly cumulative incidence proxy (passing exactly through the data points).
- The values of this spline are evaluated at daily intervals, producing a daily proxy for cumulative incidence.
- Differences of successive values of this daily cumulative incidence proxy are taken, producing a proxy for daily incidence.
And advantage of this procedure emphasized by te Beest, et al. is that the total incidence in the daily series exactly matches the total incidence in the original weekly series. When the weekly incidence series is a noisy and likely biased measure of true incidence, this does not seem to be a compelling justification.
A disadvantage of this procedure is that it can produce negative values for daily incidence. When applied to the proxies used by Kissler, et al. (what I call proxyW in Part 2), there are 6 negative values for NL63, 30 for 229E, none for OC43, and 89 for HKU1. Kissler, et al. replace these negative values with 10-7.
I’ve looked at how the other proxies I discuss in Part 2 fare in this regard. Fixing the holiday anomalies only slight improves matters — proxyWX, for example, which has a fudge for holidays, has only one fewer negative daily values than proxyW. Adjusting for outliers and zeros in the fraction of positive tests for each coronavirus has a bigger effect — proxyWXo, for example, has 2 negative values for NL63, 9 for 229E, none for OC43, and 38 for HKU1.
Negative daily incidence values are completely eliminated when the fractions of positive coronavirus tests that are for each virus are smoothed, as described in Part 2 (proxyWs and proxyAXn, for example, have no negative values).
Negative values can also be reduced by applying a filter to smooth the daily values after step 4 above. I use a filter with impulse response (0.04,0.07,0.10,0.17,0.24,0.17,0.10,0.07,0.04). After this filtering, proxyWXo has only 14 negative values (all for HKU1).
Note that it may be desirable to smooth at either or both of these stages beyond the amount needed to just eliminate negative daily incidence values, since values only slightly greater than zero may also be undesirable. I’ll show below what the effects of this smoothing are on estimates for R.
The generative interval distribution used
The formulas from above — Rt = b(t) / ∑a b(t-a)g(a) and Ru = ∑t g(t-u)Rt — involve the generative interval distribution, g(a). Estimation of g requires information on timing and source of infections that is not captured by the incidence proxy. Due to an absence of studies on the generative interval distribution for the common cold coronaviruses, Kissler, et al. used an estimate for the serial interval distribution of SARS (a betacoronavirus related to SARS-CoV-2).
The serial interval distribution is the distribution of the time from the onset of symptoms of one infected person to the onset of symptoms for another person whom they infect. If the incubation period (time from infection to developing symptoms) were the same for all people, the serial interval distribution would be the same as the generative interval distribution — the delay from infection to symptoms for the first person would be cancelled by the same delay from infection to symptoms for the person they infect. Since the incubation period is actually at least somewhat variable, the serial interval distribution will typically have a higher variance than the generative interval distribution, although their means will be the same. (See this paper for a more detailed discussion.)
In the Kissler, et al paper, using a serial interval distribution as they do is actually more appropriate than using a generative interval distribution, since the incidence proxies are not proxies for numbers of people infected each week, but rather for the number who come to the attention of the health care system each week. If people seek medical care a fixed time after developing symptoms, the serial interval distribution would be appropriate to use. (Of course, there is actually some variation in how long people wait before seeking medical care.)
The serial interval distribution used by Kissler, et al. is derived from a Weibull distribution with shape parameter 2.35 and scale parameter 9.48 (which has mean 8.4). This is a bit funny, since Weibull distributions are continuous, but a discrete distribution is needed here. Kissler, et al. just evaluate the Weibull probability density at integer points, up to the integer where the right tail probability is less than 0.01. This procedure will not generally produce a discrete probability distribution in which the probabilities sum to one. In this case, the probabilities sum to 0.995, so the error is small (it will lead to R being overestimated by about 0.5%).
The estimated SARS serial interval distribution used is from this paper by Lipsitch, et al. Here is the Weibull fit together with the count data it is based on:

The Weibull model seems fairly reasonable, except that it perhaps gives intervals of one or two days too high a probability. There is, however, no real reason to use a distribution having a simple parametric form like the Weibull — a better match with reality would probably result from just estimating these 20 probabilities, while incorporating some sort of smoothness preference, as well as whatever other biological knowledge might be available to supplement the data.
Estimates found using various proxies for incidence
Using the daily incidence values interpolated from weekly numbers, b(t), and the chosen generative/serial interval distribution, g(a), Kissler, et al. obtain daily estimates for Rt and Ru using the formulas above. Weekly estimates for these R values can then be obtained by taking the geometric average of the values for the 7 days within each week. Kissler, et al. obtain smoothed weekly values by averaging over the 21 days in each week and its preceding and following weeks.
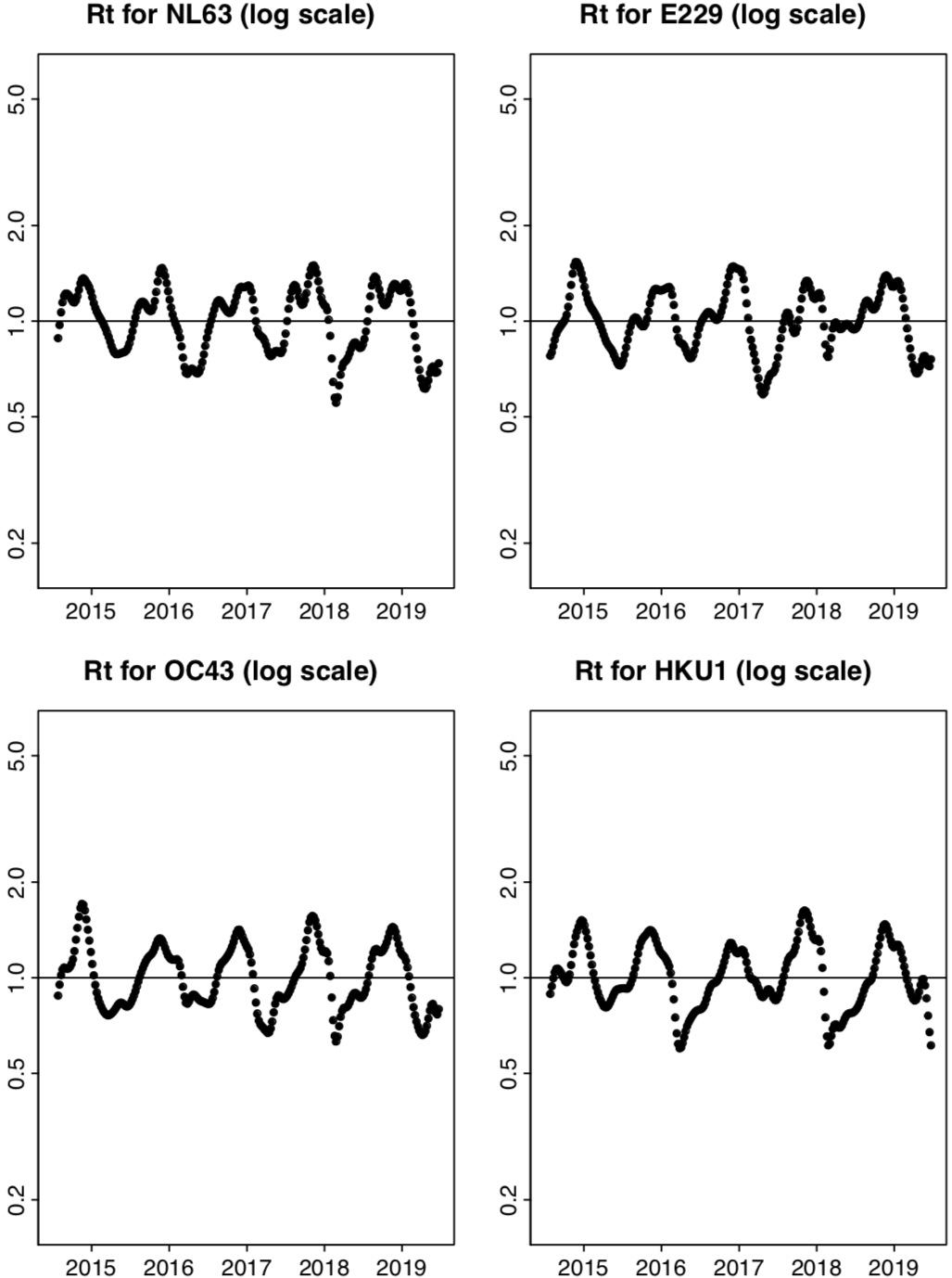
Here are the smoothed weekly values for Ru found using proxyW, the proxy used by Kissler, et al. (click for larger version):

One can see some variability that looks like it must be noise, especially for HKU1. Here are the values for Ru without smoothing:

The red points represent values that are beyond the scale of the plot.
There’s quite a bit more noise in the plots of Ru without smoothing, which may be why Kissler, et al. thought it would be beneficial to model the smoothed estimates of Ru. The benefit is illusory, however. As I’ll discuss more in the next part of this series, Kissler, et al. use a simple least-squares linear regression model for the log of these estimates. The geometric average used for smoothing Ru corresponds to a simple average for the logs. The reduction in noise from this averaging will help to no greater extent than the least-squares fitting procedure copes with noise in any case. But the averaging might obscure some relationships, and will introduce correlations in the residuals from the regression that complicate assessments of uncertainty.
I argued above that it would be better to model Rt than Ru. Here are the estimates for Rt found using proxyW:

There’s a huge amount of noise here. I’ll show in the next post that fitting a model using these Rt estimates produces ridiculous results. So if Kissler, et al. ever considered modelling Rt, one can see why they might have abandoned the idea.
Recall from above that Ru is estimated from the equation Ru = ∑t g(t-u)Rt. So the Ru estimates are weighted averages of Rt estimates. Crucially, the averaging is done on Rt itself, not its logarithm, and so has a much different effect than the averaging that is done to smooth the Ru values. In particular, some values for Rt could be zero or close to zero, without this producing any Ru values that are close to zero. This is why the estimates for Ru don’t look as bad as those for Rt.
The real problem, however, is that proxyW, used by Kissler, et al., has multiple problems, as discussed in Part 2 of this series. Better Rt estimates can be found using proxyWXss, which fixes the problem with holidays, fixes outliers in the coronavirus data, replaces zeros in the coronavirus data with suitable non-zero values, and smooths the percentages of positive tests for each virus amongst all positive coronavirus tests. Here are these Rt estimates:

These seem much more usable. Of course, the estimates for Ru and their smoothed versions also have less noise when proxyWXss is used rather than proxyW.
Using some of the other proxies discussed in Part 2 gives estimates with even less noise. Here are the Rt estimates found using proxyDn, with filtering applied to the daily proxies:

After writing Part 2, it occurred to me that by replacing the ILI proxy by its spline fit, one can create a proxy, which I call proxyEn, that is entirely based on spline fits. Such a proxy can directly produce daily values, without any need for the interpolation procedure above. Here are the Rt estimates produced using this proxy:
Future posts
In my next post, I’ll discuss the regression model that Kissler, et al. fit to their estimates of R, especially for the two betacoronaviruses that are related to SARS-CoV-2. This model has terms for immunity, cross-immunity, and seasonal effects. I will discuss problems with this model, and with how the authors interpret the results from fitting it, and look at alternatives. In subsequent posts, I’ll discuss the later parts of the paper, which model the incidence proxies in a different way using a SEIRS model, which is then extended to include SARS-CoV-2. This SEIRS model is the basis for their results that are intended to provide some policy guidance.
R-bloggers.com offers daily e-mail updates about R news and tutorials about learning R and many other topics. Click here if you're looking to post or find an R/data-science job.
Want to share your content on R-bloggers? click here if you have a blog, or here if you don't.